Article Text

Abstract
Objectives: From a series of glimpses, we perceive a seamless and richly detailed visual world. Cerebral damage, however, can destroy this illusion. In the case of Bálint's syndrome, the visual world is perceived erratically, as a series of single objects. The goal of this review is to explore a range of psychological and anatomical explanations for this striking visual disorder and to propose new directions for interpreting the findings in Bálint's syndrome and related cerebral disorders of visual processing.
Methods: Bálint's syndrome is reviewed in the light of current concepts and methodologies of vision research.
Results: The syndrome affects visual perception (causing simultanagnosia/visual disorientation) and visual control of eye and hand movement (causing ocular apraxia and optic ataxia). Although it has been generally construed as a biparietal syndrome causing an inability to see more than one object at a time, other lesions and mechanisms are also possible. Key syndrome components are dissociable and comprise a range of disturbances that overlap the hemineglect syndrome. Inouye's observations in similar cases, beginning in 1900, antedated Bálint's initial report. Because Bálint's syndrome is not common and is difficult to assess with standard clinical tools, the literature is dominated by case reports and confounded by case selection bias, non-uniform application of operational definitions, inadequate study of basic vision, poor lesion localisation, and failure to distinguish between deficits in the acute and chronic phases of recovery.
Conclusions: Studies of Bálint's syndrome have provided unique evidence on neural substrates for attention, perception, and visuomotor control. Future studies should address possible underlying psychoanatomical mechanisms at “bottom up” and “top down” levels, and should specifically consider visual working memory and attention (including object based attention) as well as systems for identification of object structure and depth from binocular stereopsis, kinetic depth, motion parallax, eye movement signals, and other cues.
- visual cortex
- visual attention
- visuomotor control
Statistics from Altmetric.com
In 1909, Reszö Bálint described in a German language periodical a striking set of visual defects in a man with bilateral hemispheric lesions.1 This Hungarian physician studied his patient from 1903 until the man died in 1906. The patient's defects were due to progressive cerebrovascular complications that began in 1894. The symptom complex included:
(1) “Spatial disorder of attention”: an inability to perceive at any one time the several items of a visual scene. This has been compared with visual disorientation (Gordon Holmes' 1918 term)2 and simultanagnosia (defined by Wolpert in 1924 as an inability to interpret the totality of a scene despite preservation of ability to apprehend individual portions of the whole).3
(2) “Psychic paralysis of gaze”: an inability to shift gaze voluntarily to objects of interest despite unrestricted eye rotations. This resembled later descriptions of spasm of fixation4,5 and acquired ocular apraxia (apraxia, Greek for “not acting”), but differed from congenital ocular motor apraxia, a childhood disorder in which head thrusts occur with voluntary refixation despite a full range of reflexive saccades.6,7
(3) Optic ataxia (ataktos, Greek for “disorderly”): difficulty reaching under visual guidance despite normal limb strength. This was not due to defective position sense (when Bálint positioned his patient's left hand, the man was able to imitate the movement with his right). Nor was it purely visual, in which case both hands should have been affected equally (the right hand was worse).
Bálint's patient showed signs of a left hemineglect syndrome: he was unaware when approached from the left or from behind, and his focus of attention was skewed 35 to 40 degrees into the right hemifield. When encouraged to look to the left, the man would see objects “immediately and always,” but attention within this area would quickly wane due to a “concentric constriction” of the “attentive field.”1 Bálint attributed the gaze disorder to the attention defect.
The patient had adequate visual acuity (20/20 in the right eye and 20/40 in the left) and visual fields (for white objects and colours) and could identify a relatively large target (such as a person) by vision alone, yet could not place a dot in the centre of a circle or triangle or perceive this error on visual inspection. Bálint suggested that his patient could “see” either the dot or the shape, but could not perceive them both at the same time. He reasoned that the man could “see” only one object at a time, regardless of the object's size.
Postmortem showed a deformed and atrophic brain, with bilateral and nearly symmetric lesions of the posterior parietal lobe, upper temporal lobe, and occipital lobe; marked damage in posterior parts of the superior and inferior parietal lobules; and lesser changes in the left central gyrus and surrounding areas. The left internal capsule was also damaged. There were no changes in the frontal lobes, optic nerve or tract, lateral geniculate body, or calcarine cortex. In a famous anatomical sketch that depicted the lateral surfaces of the patient's left and right hemispheres (fig 1⇓), Bálint emphasised the lesions in the angular gyri and the preservation of the cortex around the calcarine fissure, even though there was clearly damage in other vision related structures (including the posterior corpus callosum, bilateral white matter, upper portions of the thalami, and the pulvinar).

Bálint depicted the lesions in his patient in schematic diagrams showing the lateral surface of the hemispheres.
Gordon Holmes reported on “disturbances of visual orientation” in six soldiers with occipital wounds who misjudged the location and distance of an object, and the relative distance between two objects despite adequate visual acuity and stereoacuity.2 In 1946, Holmes explained it thus:
“As a result of visual disorientation, a patient may be able to find his way about even in familiar surroundings, and owing to the failure to recognise the positions and distances of objects, he may collide with obstacles, and may even walk into a wall though he sees it directly. He is also unable to grasp or point accurately to objects within the range of his vision.”8
The findings among these soldiers are often compared with those in Bálint's case, but also resemble the (heretofore neglected) findings in soldiers studied by Tatsuji Inouye, a Japanese ophthalmologist (see below).9 The visual field defects among soldiers studied by Holmes included homonymous hemianopia, quadrantanopia, paracentral scotomata, and extensive loss of peripheral vision. These visual field defects could not account for all the soldiers' complaints, however, because they failed to notice or attend to images that fell on retinal regions where “sensibility was intact” or “only slightly affected.”2 Holmes distinguished this problem from the unilateral disturbance of visual attention that can accompany parietal and lateral occipital lesions (the hemineglect in Bálint's). The soldiers erred in counting coins (for example, by counting the same coins twice) and had trouble navigating (that “contrasted unfavourably with a blind man”) and reaching toward targets within central or peripheral vision. Holmes attributed these difficulties to a combination of defects: the spatial localisation defect, failure to attend to objects in the periphery, and defective fixation and search. He and Horrax attributed the brain damaged soldiers' impairments in looking for and counting objects, reaching, reading, and writing to constriction of visual attention to one object (simultaneous agnosia) plus spatial disorientation.10 Holmes thought that his soldiers' inability to touch with the hand objects located in central or peripheral fields was vision related, whereas Bálint thought that the limb control defect he found was somewhat independent of a visual deficit.
Holmes estimated the survivors' brain lesions by assuming a straight course of a single missile between the entry and exit wounds in the skull. All apparently had bilateral lesions of the visual cortex in dorsal and lateral locations. A postmortem of soldier 5 showed an entry wound in the left hemisphere, destroying the supramarginal gyrus with exit from the right angular gyrus. Soldier 2 had an entry wound in the right lateral occipital lobe with exit from the left angular gyrus. Holmes thought that damage to the angular gyri and their connections could affect spatial orientation yet leave visual perception intact, and could disturb the centres from which the eye movements are elicited, with consequences for ocular fixation, accommodation for near objects, and blinking. He favoured Starling's idea that these areas performed an “integrative” function for “visual impressions” from the retina and “tactile and muscular sensations from all parts of the body.”2,10
Holmes later explained that “localisation in space by vision is not a simple perception or an innate faculty,” but “is acquired in childhood by correlating and integrating other sense impressions with visual perceptions” and depends on “integrity of anatomical paths linking visual perception with muscular and tactile impressions from all parts of the body, including the ocular musculature.” He reasserted that “prominent disturbances of visual orientation result from bilateral lesions of the parieto-occipital regions of the brain, and especially from those which involve the subcortical white matter in the neighbourhood of the angular gyri, but defective localisation in homonymous halves of the visual fields may be caused by injury of this portion of the opposite hemisphere.”8
Hécaen and Ajuriaguerra coined the term “Bálint's syndrome” in a 1954 report on four patients who exhibited sluggish exploration of their visual environment.11 The sluggishness was attributed to “sticky fixation” (restriction of fixation to a single point) and emergence of a primitive “grasp” reflex affecting the eyes. They also commented on a motor control defect due to possible damage in the posterior parietal and occipital association areas concerned with ocular pursuit. These patients had extensive brain damage, however, with impairments ranging from extensive visual loss to stupor, and therefore yielded few data to test the authors' ideas.
Luria reported on a Polish officer who was shot in the head during the second world war.12 The brain damage led to “disorders of `simultaneous perception', `vestibular giddiness', and `fits.'” Although the officer was adequately able to perceive objects, he exhibited severe constriction of the visual fields during a perimetric examination and moved his eyes between objects in a “confused and inadequate manner,” causing “failure to fixate one or more of the objects present.” Like Bálint's patient, the officer neglected objects on the left, showed defective hand movements under visual guidance, and showed similar “helplessness” of gaze.13 Luria suggested a “limitation of visual attention,” “incapacity to combine details into a coherent whole,” and “piecemeal perception” due to “weakness in cortical tonus” that caused each “focus of excitation” in the visual cortex to “inhibit the remainder of the visual cortex by negative induction.” In a famous illustration, Tyler depicted the limited ocular scan paths in a patient with Bálint's syndrome.14
Evidence suggests that Tatsuji Inouye, a Japanese ophthalmologist, made independent scientific observations that resemble and even antedate the findings in Bálint's initial report (in 1907 in Hungarian15). Inouye studied Japanese soldiers who were shot in the head during the Boxer Rebellion of 1900 and the Russo-Japanese war of 1904–5.9 (His conclusions, originally written in German, were recently made available in English.16) Inouye applied perimetry testing and used a “craniocordinometer” to estimate brain lesions from entry and exit wounds in survivors. He was familiar with the work of Henschen17 and von Monakow18 on localisation of visual function, and proposed a modern, anatomically accurate map of the primary retinotopic representation in the calcarine cortex (striate cortex or area V1) before Holmes and Lister.19 He spoke against war and advocated for soldiers' pensions based on quantitative test scores and functional outcomes.
Several of Inouye's patients (cases 5–11) had inferior hemianopia, a loss of vision in one or both of the lower visual fields, compatible with the lesions of the dorsal (parietal) visual pathways typically reported in cases such as Bálint's. Sub-lieutenant Nobuta (case 6) was wounded on 14 July 1900 in Tsien-Tsien. Perimetry (on days 55 and 824) showed bilateral inferior quadrantanopia that spared the macula. By day 61 Nobuta could avoid obstacles and by day 108 he could read individual letters, yet these did not form a unified sentence. Inouye noted a right occipitoparietal entry wound and left external occipital protuberance exit wound and marked damage in the cuneus (the dorsal visual area bounded by the calcarine and parieto-occipital fissures).
Infantryman Nambu (case 7), 26 years old, was shot in the occiput with a small calibre bullet while kneeling and shooting on 18 July 1904 at Port Arthur. He seemed confused and totally blind. He could not see in his lower visual fields and had a scotoma in the central 4 degrees of each upper field. Later, his sensorium was clear, visual acuity was 2/60, pupils, lids, and extraocular muscles worked normally, and he had no alexia, agraphia, or “psychic blindness (associative agnosia, Lissauer type20). However, he moved only with help from someone else and acted and walked as if he were blind (days 294 and 476), resembling Holmes' description. The wound was explored with temporal lobe resection. Inouye indicated that the patient had lesions of the cuneus that extended deep into the substance of the occipital pole.
Infantryman J Arai, a 25 year old (case 26), had a small calibre bullet wound to the caudal right parietal region, with exit from a symmetric location on the left. On day 75, visual acuity tested with a chart using forms was almost normal, but the doctor had to point to individual elements on the chart, otherwise Arai mixed them up. There was no associative agnosia. On day 166, while walking, Arai would sometimes bump into objects that he could apparently see (much as infantryman Nambu and Holmes' soldiers). For safety's sake Arai walked in a “crab-like fashion.” To fixate objects positioned above, below, left or right, he had to move his face in that direction, suggestive of ocular apraxia. Signs of speech disturbance were present. Visual fields showed strong concentric narrowing through day 204, but colour perception seemed to be normal. Inouye reasoned that the principal lesion affected the left angular gyrus and spared the optic radiations.
By contrast, Private Takeda (case 2) was shot in the left occiput and found lying in a bank of snow at Sha-ho on January 28, 1905, but did not show visual attentional or visual spatial disturbances as in the above soldiers or Bálint's case. Takeda had a right homonymous hemianopia that spared fixation, associative agnosia, and optic aphasia (pure alexia). He also called red green and green blue, despite the excellent lighting conditions (clear weather) under which the colour targets were presented. Inouye interpreted this as weakness of colour memory, although cerebral achromatopsia or colour anomia would be just as likely. Generally, lesions of the ventral visual association areas (along the occipitotemporal or “what' pathway), as in Takeda, tend to produce defects such as pure alexia, associative agnosia (including prosopagnosia) and achromatopsia. This differs from the pattern of defects that follow dorsal (superior occipital and parietal) lesions reported in cases of Bálint's syndrome.
ROLE OF CASE REPORTS
Historically, Bálint's syndrome has been based on single case studies. This could be because the syndrome is relatively uncommon; also, the bilateral lesions in these cases cause extensive visual, sensory, and cognitive impairments that hinder testing. Although single cases have provided valuable insights into the anatomical substrates of vision, attention, and visuomotor control, the variability of lesion effects has been underappreciated. Interesting abnormalities are likely to be reported, and less striking cases are likely to be ignored, creating a biased picture concerning lesion effects. Lesion effects and degree of recovery vary for broad reasons such as patient age, time since lesion onset, and amount of white matter involvement. Also, not all patients with similar brain lesions show the same deficits. Some patients were studied in the acute phase of injury, at the zenith of behavioural deficits, whereas others were studied in the chronic phase of recovery, months or years later. A review of two dozen reports of “simultanagnosia/Bálint's syndrome” covering the 9 decades after Bálint's initial work showed a diversity of behavioural deficits, brain lesions, vision assessment techniques, operational definition of terms, and opinions on underlying mechanisms.21 The diverse aetiologies include cerebrovascular disease, as in Bálint's original case (especially watershed infarctions),22 tumour, trauma, prion disorders such as Creutzfeldt-Jakob disease, and viral infections such as HIV. Neurodegenerative conditions such as Alzheimer's disease23,24 might be the most common cause of Bálint's syndrome.
Scientific concerns surround key cases upon which Bálint's syndrome is grounded, due to failure to assess basic visual functions (other than acuity and, occasionally, stereopsis), and to consider confounding eye conditions (such as retinopathy, cataracts, or optic neuropathy). In this vein, Bálint1 claimed that his patient had normal visual fields but it is unclear which procedures support this conclusion in a man who was likely difficult to test. It is risky to generalise from historical or modern single case reports in Bálint's syndrome.
SIMULTANAGNOSIA VERSUS VISUAL DISORIENTATION
Key case studies on which Bálint's syndrome is based have proposed differing mechanisms to account for the perceptual underpinnings of the disorder. Bálint suggested that his patient's main problem was a spatial disorder of attention. This occurred in the context of a left visual hemineglect syndrome. Bálint inferred that his patient could see only one object at a time no matter what size, a condition later called simultanagnosia.12 Yet, Bálint's findings indicate that his patient could actually process multiple stimuli in some circumstances (for example, while navigating).25 Also, object recognition can proceed economically from just a few key features, and identifying a person or object is not the same as seeing him, her, or it all at once. Along these lines, Holmes explained that “recognition does not depend on synthesis of all the sense impressions derived from an object, the object is usually recognised as a whole, not as the sum of its parts and it is not necessary that the mental image should correspond exactly with the perception of the new object, or contain representations of all its features; images stored in memory are particulate, that is, they consist of a number of parts each of which represents a feature of the object remembered.”8
Simultanagnosia should not be confused with agnosia, an associative (or mnestic) disorder, in which percepts are “stripped” of their meanings.26 In Holmes' words, agnosia is the “inability to recognise objects perceived by one of the senses” that occurs “when sense impressions are unimpaired and mental images are intact.”8 Patients with agnosia can no longer recognise previously familiar objects or learn new objects; in a restrictive form, prosopagnosia, the defect is most apparent for faces.22 Patients with “Bálint's syndrome” may not recognise objects due to their perceptual impairments, as described under Lissauer's old term “apperceptive agnosia.”19 In reality, most people with visual object recognition defects probably have a combination of associative and perceptual deficits.
Wolpert's definition of simultanagnosia can be operationalised as an inability to report all the items and relations in a complex visual display, despite unrestricted head and eye movements. A suitable screening tool is a picture containing a balance of information among the four quadrants, such as the Cookie Theft picture.27 The patient's report can be correlated with a checklist of the items in the picture. Exclusion criteria should include aphasia severe enough to impair the verbal descriptions of a display, to avoid confusing a defect of language with one of visual perception. It is also crucial to be aware of defective visual acuity or fields. Extensive visual field loss caused by double homonymous hemianopia (or “keyhole vision”) (see the case of Luria12), or a central or paracentral scotoma, may hinder simultaneous perception and visual search, and objects may even seem to vanish.28 Rafal commented that Wolpert's definition of simultanagnosia is specific to figure drawings but is more general than the constriction of attention seen in Bálint's syndrome.29 On the other hand, simultanagnosia has been reported with bilateral lesions of the superior (dorsal) portions of both occipital lobes in Brodmann's areas 18 and 1928 in patients who do not display the full picture of Bálint's syndrome seen after lesions of parieto-occipital areas and angular gyrus,29 suggesting that simultanagnosia may arise from damage to different areas of an interconnected network of areas responsible for perception and attention (fig 2⇓).

Transverse CT images of the brain in a patient with simultanagnosia show bilateral lesions restricted to the superior visual association cortex of the occipital lobes (visible as dark or wedge shaped areas located at the bottom portion of each image).28
Holmes2 and Holmes and Horrax10 distinguished between visual disorientation and the unilateral defect of visual attention and the defect of simultaneous perception emphasised by Bálint. Visual disorientation was due to errors in judging the location and distance of an object, and in judging the relative distance between two objects. Although Bálint's syndrome depends on both visual spatial disorientation defects and the constriction of attention,29 the inability to see more than one object at a time due to simultanagnosia seems to be sufficient to account for difficulty judging the distance between two objects, a crux of visual spatial disorientation.2,10
As we shall see, visual disorientation (as in Holmes' soldiers) and simultanagnosia (in cases such as Bálint's) might each be generated out of differing combinations from the same large menu of defects. This includes low level spatial resolution problems (affecting visual acuity and contrast sensitivity), visual field defects (such as tunnel vision and paracentral or central scotomata); motion processing deficits (cerebral akinetopsia), impaired ocular pursuit of moving targets, and impairments of visual attention and visual working memory. There are multiple cues to structure and depth that could be affected by brain lesions that cause Bálint's syndrome, yet the role of these potential defects in the generation of Bálint's syndrome is virtually untested.
SYNDROME STATUS
Bálint's syndrome may not exist as a sufficiently autonomous complex to satisfy Benton's criteria30 for a syndrome.25 Firstly, the individual components of Bálint's syndrome are not as closely bound as might be expected. For example, simultanagnosia is doubly dissociable from optic ataxia.22 Ocular apraxia does not occur independently of visuospatial defects, but is itself often absent. Secondly, the individual components of Bálint's syndrome (such as simultanagnosia and optic ataxia) seem to represent relatively broad categories comprising other more specific defects. Thirdly, patients often have confounding behavioural defects, as in the cases of Hécaen and Ajuriaguerra.11 Finally, the terminology and related operational definitions of syndrome components vary across studies.
Bálint's syndrome has been construed as a biparietal syndrome,29,31 yet does not seem to have the specific neuroanatomical significance (of damage in the angular gyri) originally attached to it and may occur with other combinations of lesions. Consider a lesion of the left lateral geniculate body or optic radiation causing right homonymous hemianopia, followed by a lesion in the right parietal lobe producing left hemineglect. Vision might be restricted to the neglected field and the patient would seem to have Bálint's syndrome (see the comparison below with hemineglect syndrome). Also, it is not clear that the components of Bálint's syndrome have any special status compared with other defects associated with lesions in a similar location. Lesions of the right angular gyrus often correlate with hemineglect, and lesions of the left with aphasia and Gerstmann's syndrome (agraphia, acalculia, and right/left disorientation).30 Bilateral lesions around the angular gyri have been associated with a profound defect of visual motion processing,32 a defect not emphasised by Bálint.
Holmes recognised that visual disorientation was strongly associated with multiple behavioural deficits:
“As a result of visual disorientation, a patient may be able to find his way about even in familiar surroundings, and owing to the failure to recognise the positions and distances of objects, he may collide with obstacles, and may even walk into a wall although he sees it directly. He is also unable to grasp or point accurately to objects within the range of his vision . . .Topographical memory is often disturbed, too; the patient is then unable to visualise or describe routes with which he is familiar, as how to go from one room to another of his house, or from his home to a neighbouring shop or station. Occasionally the distinction of right and left of his own body or of spaces is lost; this may complicate acts in which visual guidance is required . . .Visual memory, or ability to visualise familiar persons or objects, may also be defective . . .Visual agnosia is often combined with spatial disorientation or spatial agnosia, or with defects of topographical memory or imagery, and sometimes with inability to describe familiar objects which are known chiefly by their visual.”8
Bálint's syndrome may occur with bifrontal lesions33 and, perhaps, with pulvinar lesions.34 Lesions of Brodmann's areas 6 and 8 (the frontal eye fields) may hinder voluntary saccades, visual search, and scanning, compatible with ocular apraxia. Elements of optic ataxia are associated with a wide variety of lesions in Brodmann's areas 5, 7, 19, 39, 37, and a profound reaching disturbance has even been reported with a relatively mesial right temporo-occipital lesion.35 Optic ataxia may comprise a family of defects. Inability to reach and grasp targets in these cases is often multifactorial, including V1-type visual field defect, defective visual attention, and inability to locate targets with the eyes. Another possibility (described further below) is abnormal sensorimotor transformations, an inability to transform the visual coordinates of external objects to appropriate limb coordinates for generating accurate reaches36 (see the section below: Visually guided reaching and optic ataxia).
DORSAL SIMULTANAGNOSIA VERSUS VENTRAL SIMULTANAGNOSIA
The primate visual system is partly understood in terms of parallel pathways beginning in the retina. The parvocellular or P pathway, named for its connections to the simian striate cortex (area V1) via parvocellular layers 3 to 6 of the lateral geniculate body, is characterised by colour opponency and slow conducting axons that convey sustained signals.37,38 This “ventral” or temporal (“what”) pathway has strong projections to secondary areas such as V4, V8, and IT (inferotemporal area), located in the inferior occipital lobe and adjacent occipitotemporal regions.39 Damage in these regions may impair pattern recognition and learning, producing agnosia for objects and faces (prosopagnosia) and inability to read despite prior literacy (alexia). Also, damage to these areas can reduce colour perception in the contralateral field, a condition known as cerebral achromatopsia (see Inouye's description above of private Takeda 16).40,41
By contrast, the magnocellular or M pathway is comprised of large diameter, fast conducting axons that convey information about transient visual signals, connects via V1 to secondary visual areas including area V5 (mediotemporal area) lying along the “dorsal” or parietal cortical (“where”) pathway, and is thought to support motion perception. Damage in these areas produces disorders of spatial-temporal analysis (the inability to judge object location, distance, orientation, size, or motion) as well as marked disturbances of visually guided eye and hand control. The hemineglect syndrome and Bálint's syndrome are remarkable examples.29
Against this backdrop, it has been suggested that patients with Bálint's syndrome have “dorsal simultanagnosia” with attentional limitations that preclude the detection of multiple objects, whereas patients with “ventral simultanagnosia” have left occipitotemporal lesions with slowed visual processing speed, causing a difficulty in simultaneously recognising the individual parts of a multipart object, with manifestations such as letter by letter reading.42–,44 Proposing different forms of simultanagnosia reinforces the myriad behavioural differences between patients with dorsal and ventral system lesions, which have been well described (as in the literature on acquired alexia in patients with left occipitotemporal lesions45). Although patients with ventral lesions and those with dorsal lesions may both seem to have simultanagnosia, the underlying causes of the simultanagnosia are different depending on the location of the lesion (dorsal versus ventral). Further research is necessary, however, on these two categories of simultanagnosia.25 The dorsal and ventral systems might conceivably share similar processing mechanisms, which could allow a dorsally damaged patient to present with behavioural impairments characteristic of ventral damage. For example, slowed processing of letter patterns can occur with lesions in dorsal areas such as the angular gyrus and might make such a patient seem to read in a letter by letter manner. Thus, generalised slowing can be caused by lesions in either dorsal or ventral visual areas and can help to explain information processing impairments and performance loss at multiple levels46 on cognitive tasks ranging from simple memory to complex reasoning and spatial abilities.46,47 This includes reduction of the useful field of view, a constriction of the visual fields in the absence of visual sensory loss on standard perimetry testing.48,49
Of note, visual mechanisms for object recognition and localisation seem to depend on processing of signals in structures that are outside the visual cortices proper, such as the frontal lobe50,51 and cerebellum.52–,55 We should consider the potential importance of lesions in these areas to form a more complete picture of Bálint's syndrome. The prefrontal cortex also seems to be a destination of visual signal processing that begins in the primary visual cortex and plays a part in visual attention by altering the bias among competing signals in the extrastriate cortex arising from different parts of the visual field.51 Ventral pathways for object vision seem to connect with ventrolateral prefrontal areas involved in working memory for objects, whereas dorsal pathways involved in visuospatial processing connect with dorsolateral prefrontal areas involved mainly in working memory for spatial locations. The prefrontal cortex also seems to support mechanisms for visual working memory, the process of maintaining a representation of information for a brief period of time so that it is available for use.50 Impairments of working memory for objects and their locations could potentially contribute to perceptual manifestations reported in Bálint's syndrome.
Patients who meet operational criteria for Bálint's syndrome can also have damage to several of these mechanisms for visual working memory, as in the case of Bálint's patient, and the well studied modern day case of patient RM.31 RM's lesions occupied the parieto-occipital areas and white matter about the angular gyrus and posterior superior temporal lobe (see Friedman-Hill et al,31 their figure 1⇑), but also affected the left supplementary motor area and both cerebellar hemispheres. The cerebellum, with its multiple visual and motor connections,53,54 may support neural mechanisms that distinguish between image movement across the retina and self movement, and may contribute to perception of form from temporal structure alone (with the transient system carrying the signal via dorsal visual pathways).56
ROLE OF ATTENTION AND WORKING MEMORY
Visual cues convey a wealth of information about physical objects and their relations. Conscious appreciation of this information is therefore likely to require focused attention. Many images compete for our attention, yet we cannot appreciate more than a few items at once. We experience items that we attend to for whatever specific purposes we have in mind, and we fail to perceive the rest.57,58 Without focused attention, the obvious may be hard to see: we can be oblivious to marked changes in scenery (“change blindness”), and traces of retinal images in visual short term memory (visual working memory) will fade without being consciously perceived or remembered (“inattentional amnesia”).59 The very act of perceiving one item in a rapid series of images momentarily inhibits the ability to perceive another image (the “attentional blink”). In short, we seem to construct our reality from a sequence of glimpses, and our impression of a seamless and richly detailed visual world is something of an illusion.57 Damage to visual cortex and connections can destroy this illusion, resulting in a piecemeal experience in which the visual world seems to be constricted to an erratic focus containing a single object at a time, as in the symptom complex described by Bálint.1 Evidence from patients who display elements of this syndrome is relevant to the understanding of the neural substrates of visual perception, motor control, short term (working) memory, attention, and even consciousness.
Bálint's report provided early information that the visual cortex and connections among visual areas play a crucial role in visual attention. This role is carried out at several functional levels and seems to depend on both dorsally and ventrally located visual cortices and their connections. Objects in the visual field always compete for focal attention.60–,62 Although visual attention may amplify extrastriate neuronal responses to a stimulus at one spatial location in the visual field, recordings in the extrastriate cortex of monkeys suggest that such effects rely on competitive interactions among neurons representing all stimuli in the visual field.51 Attention can be biased toward target stimuli by spatial and non-spatial processing as well as bottom up and top down processing, suppressing neuronal representations of behaviourally irrelevant stimuli in the extrastriate cortex.51,63 Top down influences may derive principally from neuronal systems for working memory.51 Patients with Bálint's syndrome might fail at both top down and bottom up levels, as suggested by PET studies of visuospatial attention.64 The inability to perceive more than one object at a time in Bálint's syndrome suggests the importance of a parietal based spatial attention network in recognising multiple objects, consistent with computational models of attention and recognition.65,66
Activation of frontal areas during interactions between visual processing, working memory, and sustained attention, derive from PET, fMRI, magnetoncephalography, and event related potential research.67 The prefrontal cortex is involved in behavioural tasks that require decisions based on working memory, and visually related areas of the prefrontal cortex would represent the “final point” in a proposed hierarchical sequence of visual signal processing that begins in the primary visual cortex.68 Ventral pathways for object vision seem to connect with ventrolateral prefrontal areas involved in working memory for objects. Dorsal pathways involved in visuospatial processing and generally implicated in Bálint's syndrome seem to connect with dorsolateral prefrontal areas involved mainly in working memory for spatial locations.50 Recognition of a specific visual target among equally familiar distracters is associated with both an enhanced response to that target in the frontal cortex that persists with target repetition, and a reduced response in the extrastriate visual cortex with stimulus (target or distracter) repetition, a process that may fail in the case of Bálint's syndrome.69 These complementary neural mechanisms track the status of familiar items in visual working memory, allowing efficient recognition of a relevant object and rejection of irrelevant distracters.70 Failure of these mechanisms in neurological conditions such as Bálint's syndrome can provide information about how (1) spatial attention provides the “glue” that allows elementary features such as colour, shape, or size belonging to a stimulus to be grouped (the “binding problem”); (2) image features such as line segments are grouped to form larger structures such as surfaces or regions; (3) object parts are encoded relative to a reference point on the object itself (an object centered representation); and (4) focal visual attention interacts with visual features such as colour and shape to guide visual target search.71
In “biased competition” accounts of visual search,51,63,72 attentional selection is guided by: (1) bottom-up sources that arise from perceptual processes and (2) top-down sources that arise from goal directed, executive processes. Lesion studies illustrate the role of the visual cortex in maintaining the perceptual processes that operate on the bottom up inputs in the biased competition model and also are compatible with results in non-human primates, indicating the role of both V4 and V5 (mediotemporal area) in bottom up attentional processing. For example, Schiller73 maintains that simian V4 is part of a neural circuitry that contributes to the selection of crucial, physically less prominent stimuli, which in early parts of the visual system evoke less activity than other stimuli in the visual scene. In addition, the receptive fields of neurons in V4 and IT are suppressed when attention is directed to one of two targets, as if the receptive fields were shrinking around the attended location.74 This resembles shrinkage in the useful field of view measured by patients with Bálint's syndrome and with lesions of the early visual cortex.75 Shrinkage of the useful field of view due to reduced processing speed and attention rather than hemianopia has since been identified as a consequence of aging48,49 and age related neurological impairments.75,76
The visual fields in Bálint's patient were easily fatiguable, resembling the effects of fatigue in normal observers (such as air traffic controllers) with prolonged and intensive monitoring of visual displays.77,78 Holmes noted such effects in measuring the fields of vision in the clinic and wrote: “Even in normal persons the field becomes smaller and the reply is less accurate as the subject tires; it is therefore often advisable to interrupt the examination for a time, or rest the subject, if there is any sign that his attention is flagging.”8 The effects in Bálint's patient, however, were pathological and did not require prolonged effort. Still, Bálint's patient could “immediately and always” “see” targets in his neglected left hemifield with the aid of verbal prompts, suggesting that Bálint's patient could “disengage” attention from the right visual field, “shift” it to the left, and “engage” targets there like normal subjects79,80 and some modern lesion cases.81,82 In other cases, however, disengagement of attention may be abnormal, and ocular apraxia and simultanagnosia might even be explained by an inability to disengage attention from fixated objects,42,83 reminiscent of the visual grasp reflex hypothesised by Hécaen and Ajuriaguerra in their cases.11 Patients may have to close their eyes to break fixation from one object so they can look at another. If attention is needed for conscious perception of objects, restriction of attention to the fixated object might help explain simultanagnosia.
Bálint's syndrome might also reflect loss of a master spatial map within a feature integration framework.84 In this framework, attention operates beneath conscious awareness, helping the mind to group different features and attributes of objects. Patients with cerebral lesions may be unable to identify which features in the panorama (for example, colour and shape) belong to an object and may therefore form “illusory conjunctions.”31 For example, when presented with a rapid presentation containing a red “O” and a green “X”, the patient might erroneously perceive a green “O” and a red “X”. In a similar vein,29 when patients with Bálint's syndrome view overlapping figures (such as a comb and a spoon), they see one or the other, but not both; alternatively, they may see neither and therefore generate an incorrect answer by focusing on local elements in the display (analogous to the parable of the blind man and the elephant).
Patients with Bálint's syndrome may have difficulty switching attention between local details and global structure—that is, they cannot see the forest for the trees. This type of deficit can be investigated using hierarchical stimuli such as a large “global” letter made of smaller “local” letters (for instance, a large letter “C” made of multiple smaller versions of the letter “s”). Neural mechanisms of global/local attention may differ from mechanisms for object/part attention. Attention to objects and parts can be tested using objects having three parts: a vertical body and two crossbars (fig 3⇓).85 There are two features (small bumps) at the end of the crossbars. These bumps can be on the same crossbar or on different crossbars. Subjects are asked to compare the bumps and report if they are identical or different. Normally subjects are better able to compare features that are on the same part (the same crossbar) than those on different parts (different crossbars), but these results may vary in patients with neurological impairment such as Bálint's syndrome. If part attention and spatial attention are controlled by separate neural processes, patients with ventral pathway damage should not show part attention effects (better performance in the same-part condition than in the different-part condition), but should show intact spatial attention (better performance when bumps are near one another regardless of the part they are on). It is possible that subjects with damage to the dorsal visual pathway (as in hemineglect) might show the opposite pattern; that is, intact part attention but disrupted spatial attention. For example, in hemineglect, a part that crosses the visual midline may reduce hemispatial neglect or extinction, consistent with some reports of “object centred” neglect.86 Further, patients with Bálint's syndrome might exhibit a simultanagnosia for multipart objects, failing to appreciate all of the parts simultaneously and only reporting one of the parts.

Examples of stimuli used to study part based attentional selection.71 Observers are asked to judge if the critical features (the “bumps”) of an object look the same or different. The bumps can appear on either the same part or on different parts, but are always located the same spatial distance from one another. Neurologically intact observers are faster to discriminate critical features that are located on the same part of an object.
Experiments using the Sternberg paradigm87 can reveal connections between basic visual processing, memory, and attention in patients with cerebral disorders of vision such as Bálint's syndrome.88 Subjects view a series of stimuli (complex patterns that resist verbal encoding) and judge whether a subsequent probe stimulus was part of the series. Performance varies with series length and position of the probe in the series, more recent positions allowing better recognition. When the stimulus series contains a single item, the procedure comprises a simple visual discrimination paradigm, but with additional items in the series, it constitutes a recognition paradigm. Varying the dimensions of stimulus similarity permits quantitative assay of recognition memory's potential stimulus related and task dependent antagonists, including both visual and memory factors. Visual recognition memory uses different neural pathways, depending on stimulus attributes, task demands, and behavioural goals.89 This memory is robust over short time periods for basic attributes such as spatial frequency, orientation, and motion direction,90 allowing normal subjects to compare and make inferences from relations between different items in the environment, but declining rapidly with interference from successive inputs. Sternberg's recognition memory procedure87 can be used to examine the effects of such interference on the visual system.88 Results in normal subjects show that series length effects in recognition memory reflect both the number of intervening items and the length of the number of items in the series. Recognition is unaffected by delays of up to 4 seconds. Time alone is not a major source of forgetting with short intervals, even with complex stimuli. However, performance may decline with lesions in visual pathways, or with deficits of visual working memory that depend on prefrontal cortex.50 Alzheimer's disease affects vision91,92 and visual attention76 and may cause Bálint's syndrome. Pilot studies in our laboratory in four persons with Alzheimer's disease, conducted in collaboration with Dr Robert Sekuler, showed that the results with series length (n=2) may help distinguish between impairments of early vision and visual working memory and attention.
What are anatomical substrates of the attentional mechanisms that may be impaired in Bálint's syndrome? Current evidence suggests the following hypotheses:
(1) Visuospatial attention and temporal attention depend on a network of structures that includes occipital areas 17, 18, and 19 (both dorsal and ventral), connections with temporal and parietal lobe regions (especially the inferior parietal lobule on the right), frontal eye fields, and prefrontal cortices.
(2) The ventral pathway is crucial for bottom up object representation and memory.
(3) The ability to perceive objects in rapid serial visual presentations depends on dorsal (psychophysical transient) pathways and their parietal connections.
(4) Executive switching of attention between visual objects and tasks depends on areas in the prefrontal cortex that influence the dorsal and ventral visual pathways in a top down manner.
Bálint's syndrome and the hemineglect syndrome probably affect several of these hypothetical mechanisms, including spatial attention, the enhancement of object representations, and the encoding of objects into visual working memory.
RELATION BETWEEN BÁLINT'S SYNDROME AND THE HEMINEGLECT SYNDROME
Elements of the hemineglect syndrome, a disorder of attention in which patients with right hemispheric lesions (usually) fail to register targets in the opposite panorama, feature prominently in reports by Bálint and other influential researchers; a diagnosis of hemineglect has never excluded a diagnosis of Bálint's syndrome. It is possible that Bálint's syndrome is distinct from hemineglect syndrome because the critical lesions in Bálint's syndrome are bilateral and occupy the parieto-occipital junction, whereas lesions in the hemineglect syndrome occupy the temporoparietal junction and typically affect the angular gyrus.29 Yet these anatomical distinctions are not clear cut, and the behavioural overlap between these two syndromes is obvious.
Patients with hemineglect syndrome may have defective visual search (using eye movements93 that resemble ocular apraxia) and defective hand movements under visual guidance (with both hands into the neglected hemifield and with the limb on the neglected hemibody to all fields) that resemble the pattern of optic ataxia.94,95 Patients with hemineglect have visuoperceptive disturbances that resemble simultanagnosia, including “extinction” of objects in the “bad” hemifield when stimuli are present in both fields, or even when multiple stimuli are simultaneously displayed in the “good” field alone.
Bálint's patient had left hemineglect due to extensive damage in the right inferior and superior parietal lobules. Lesions in these locations are likely to produce visuomotor difficulty in reaches conducted with the left hand to both visual fields (“hand effects”) and with both hands to the left visual fields (“field effects”).96,97 Bálint emphasised that his patient had trouble locating visual targets with the right hand, but left hand control could also have been impaired. Observations on the reaching behaviour in patients with focal circumscribed lesions of visual cortex can help clarify these issues.
Holmes8 commented on “attention hemianopia” affecting one or both hemifields. He wrote that “when a subject's attention is poor, or when it tires, his replies to stimuli, especially those falling in the periphery or less sensitive portions of the retina, tend to be inaccurate and unreliable. Under these conditions, the visual fields, particularly when mapped up by a perimeter, become gradually smaller and the contours irregular, this is frequently so in hysteria and in cases of exhaustion.” He also noted that a “specific local loss of visual attention in the hemianopic halves of the visual fields, or in some portions of them, may, however, result from cerebral lesions involving the opposite parieto-occipital lobe. When it is present the field may be normal and isolated stimuli perceived with accuracy, but no reponse may be obtained, or only a proportion of the stimuli are perceived, on the affected side which attention is diverted or claimed by another stimulus. This can be demonstrated by the observer holding up his two hands, one on each side of the fixation point and at roughly equal angles from it, and requesting the patient to reply to every movement of a finger or thumb. Regular responses are obtained if a finger on either side only is moved, but on simultaneous movements of the fingers to both the right and left, that on the affected side is not perceived or a proportion of them only may be noticed. The intensity of the stimulus to the affected side has little influence on this phenomenon; it may, for instance, be demonstrated by using a test object one centimeter in diameter on the normal side and one of 20 centimeters on the other.” He further states that “when vision is reduced in the homonymous halves of the field the responses to small objects may be irregular and inconstant; it is therefore necessary to exclude visual defects and use large stimuli in testing for visual inattention.”8
Riddoch reported that “spatial disorientation may occur in homonymous half fields alone.”98 He studied two patients who had “unmistakable” visual disorientation in the right homonymous hemifields. Both patients were unaware of the defect, which persisted until death. Both died several months after a neurosurgical procedure that showed a malignant tumour of the left parietal lobe. Visual acuity was reduced slightly to moderately. The first patient, a 58 year old retired businessman, had enlargement of the blindspot in his left eye, probably due to increased intracranial pressure, but had no hemianopic defect. This patient could quickly fix his gaze on an object and maintain it to his left, but was less consistent to his right, which resembles the defect in hemineglect. The second patient, a 34 year old farmer, had papilloedema and an incomplete right homonymous hemianopia with gross defect of localisation of objects and estimation of distance in his right hemifields. He usually pointed below and beyond objects. Visual attention was impaired in the right half fields. Although visual fields and attention improved after surgery, the patient still showed “gross spatial disorientation, especially for absolute distance.” In his defective fields, however, “he recognised, without hesitation, the shape of a cube, a sphere, a pyramid, and a cylinder.” Riddoch downplayed the role of a visual attention defect, in line with “Holmes' opinion that it does not form an essential part of the syndrome of disordered space perception.” Riddoch concluded that (1) visual disorientation can result in the fields opposite a parietal lobe lesion; (2) the defect can be dissociated from other related disorders of visual function that follow lesions in roughly the same part of the brain; and (3) the defect is not due to homonymous hemianopia or poor visual resolution. He cautioned that because a patient could be unaware of the deficiency, it may not give rise to obvious disability and must therefore be sought by the examiner. He felt that he had “nothing to add to the views already expressed by Holmes on the mechanism of the disturbance” (visual disorientation) and agreed that “so far as is at present known, the nodal point seems to be in the region of the supramarginal and angular gyri in the parietal lobes.”98
A neglected hemifield processes information much as a simultanagnosic field does. Patients with hemineglect syndrome may have difficulty making feature conjunctions in the aberrant field and may make “illusory conjunctions” (for example, of colours and letter shapes, as described above).99 Likewise, Bálint's syndrome patient RM reportedly miscombined colours and shapes, even under free viewing conditions, possibly due to an inadequate early spatial representation or map of object features.31 Diminished ability to bind features for unified perceptual objects represents yet another similarity between the hemineglect syndrome and Bálint's syndrome. RM reportedly had “spatial disorientation, optic ataxia, psychic paralysis of gaze, hyperattention to a single object, and difficulty reporting more than one object in natural scenes due to embolic strokes affecting both parietal lobes. Like Holmes' patients, RM could not judge accurately whether an object was moved toward or away from him. Visual acuity, spatial contrast sensitivity, colour, and stereopsis were adequate. Goldmann fields showed an inferior nasal depression in the left eye only, and there was no reported evidence of hemineglect. It was not stated whether RM still met operational criteria for Bálint's syndrome about 2 years after syndrome onset, when his ability to make feature conjunctions was tested. Patients with Bálint's syndrome, similarly to those with hemineglect syndrome, can recover dramatically after initial symptoms resolve.
Visuoperceptual deficits in the hemineglect syndrome, as in Bálint's syndrome, are not limited to one hemifield. One source of supportive evidence comes from studies of the “attentional blink”, which refers to the inability to identify objects in the aftermath of visual recognition (fig 4⇓).100–,102 This attentional blink normally lasts about 0.5 seconds and is thought to depend on the ability to allocate attention over time and consolidate representations in visual working memory101,103; it may also involve scene representation,104,105 postperceptual processing,103,106 and magnocellular visual pathway function.107 Husain et al tested the attentional blink in eight subjects about 1 month after a right hemispheric stroke affecting the inferior parietal lobe, inferior frontal lobe, or basal ganglia.108 These patients had visual neglect defined clinically and on a shape cancellation task, but also had trouble perceiving foveal targets in a rapid serial visual presentation task that used letter shapes as visual targets. They could not identify a second foveal target until 1440 ms had elapsed after identifying the first target, indicating a pathologically increased attentional blink. They concluded that “visual neglect is an inability to direct attention in time, as well as space.”108 Increased attentional blink is not specific to the hemineglect syndrome109 and similar limitations may arise out of simultanagnosia or Bálint's syndrome,31,108 dementia,76 or cognitive aging.110 The increased AB that follows damage to the dorsal pathway could be due to damage to an attentional network that does not direct attention in time itself, but rather provides input to a visual working memory system. The degraded inputs to visual working memory that arise from a damaged parietal based attention system could impair visual working memory, leading to an increased attentional blink.

Schematic representation of stimuli used in a rapid serial visual presentation task (A and B) in studies of the attentional blink.109
PERCEPTION OF STRUCTURE AND DEPTH
Theories of visual perception include structuralism, gestaltism, and ecological optics, with varying emphases on nativism (vision is innate) versus empiricism (vision is acquired through experience), atomism (a visual field is the sum of its parts) versus holism (the appearance of one part is influenced by the presence of others), organism (perception depends on structure of the organism) versus environment (perception depends on external stimuli), and underlying principal analogy (for example, to chemistry, physical field theory, or mechanical resonance111). Yet another varying emphasis regards method of research. For example, trained introspection versus naïve introspection (emphasised by structuralists) versus stimulus analysis (emphasised in ecological optics by Gibson112 and later by Marr 113). Bálint primarily relied on a descriptive approach and introspective analyses. The approaches of Holmes and Inouye were more in line with current strategies, which favour a combined cognitive neuroscience/visual psychophysics approach, with emphasis on optics and stimulus analysis.
The various perceptual phenomena reported in cases such as Bálint's patient (including “vanishing” objects, tilted vision, metamorphopsia, and palinopsia) are not likely to be the behavioural expression of a single mechanism (such as the inability to see more than one object at a time25), but may be better explained as a variety of combined deficits from lesions of the dorsolateral visual association cortices, which includes the putative human mediotemporal area complex and its projections to the parieto-occipital cortex. Damage to these areas and to the cortices surrounding the angular gyrus and parietal insular cortex can disturb multiple aspects of attention, as well as spatial and temporal processing, including the perception of visual motion,32 structure from motion and dynamic stereopsis, the perception of egomotion, and coordination of visual (eye centred) and vestibular (gravity centred) coordinate systems that orient us in the physical world. Bilateral lower quadrantanopias may co-occur (as in Inouye's and Holmes' soldiers) from damage to the visual cortex along the dorsal banks of the calcarine fissure, worsening a patient's overall problem.
Impairments in the perception of objects and the relation between objects that contribute to Bálint's syndrome may be related to lesions in neural structures that support perception of structure from motion and motion parallax and related processes. Holmes touched on these mechanisms when he commented that “localisation in the coronal plane depends mainly on the local signs of the retinal point excited by the image of the object, but the estimation of distance is a more complicated process which requires integrations of impressions of various origins, as those provided by binocular vision, proprioceptive impulses from the ocular muscles (especially those excited by convergence and accommodation), and judgments based on distinctness, light intensity, and the comparative size of familiar objects. Stereoscopic vision, which is related to the estimation of distance, may be affected at the same time.”8
Indeed, there are many cues to structure and depth that could be affected by brain lesions in Bálint's syndrome. Among these information sources on object structure and depth are accommodation, convergence, binocular disparity, motion parallax, texture accretion/deletion, convergence of parallels, position relative to horizon, relative size, familiar size, texture gradients, edge interpretation, shading and shadows, and aerial perspective.113 These clues are optical (except accommodation and convergence, which are ocular), monocularly available (except convergence and binocular disparity),statically available (except motion parallax and texture accretion/deletion, which are dynamic) and relative (except accommodation, convergence, and familiar size, which are absolute). Most of these information sources are quantitative (except texture accretion/deletion, edge interpretation, shading and shadows, and aerial perspective, which are qualitative).111
PERCEPTION OF STRUCTURE FROM MOTION
Perception of structure from motion or kinetic depth is a long hypothesised real world use of motion perception111,114 that may fail in patients with lesions in the dorsolateral visual cortices. Structure from motion is efficient compared with other cues such as colour and luminance.115 Models of structure from motion extract discontinuity boundaries in a field of local motion vectors, with grouping based on common direction or speed.116 Schiller73 found that mediotemporal area lesions, along with V4 lesions, produced some deficit in the perception of structure from motion. Marcar et al117 found that mediotemporal area neurons in macaque responded to kinetic boundaries, but that this processing was particular for the change in local motion at the boundary and did not code the actual orientation of the boundary. Instead, neurons in the inferotemporal cortex show orientation selectivity for shape from motion as well as shape from luminance cues. Additionally, using functional imaging techniques, Orban et al118 found a right hemispheric area near but separate from the putative location of the human mediotemporal area homologue that was selectively active during a motion defined form task. The lesions reported in cases of Bálint's syndrome often include these areas.
For an objective test of structure from motion, we developed a two AFC shape identification task that asks a subject to report the shape of the object presented in each trial (fig 5⇓). Accurate performance depends on the subject's perception of the figure's shape from motion. To prevent shape identification from non-motion cues, such as edges or dot density, and to index the difficulty of the task, varying amounts of random dot noise are added to a background region surrounding the target. The perception of structure from motion was impaired in early Alzheimer's disease70 and “motion blind”(akinetopsic) subject LM.32 LM's ability to perceive structure from motion broke down at moderate levels of moving and stationary noise.119 Aspects of LM's complaints, namely that she would see objects in one place or another but not moving in between,32 are reminiscent of complaints in Bálint's syndrome. However, LM's complaints depended on moving stimuli whereas the complaints of Bálint's patient did not. Relevant to instrumental activities of daily life, structure from motion deficits in subjects with brain lesions are associated with increased relative risk for safety errors and car crashes in high fidelity driving simulation scenarios.120
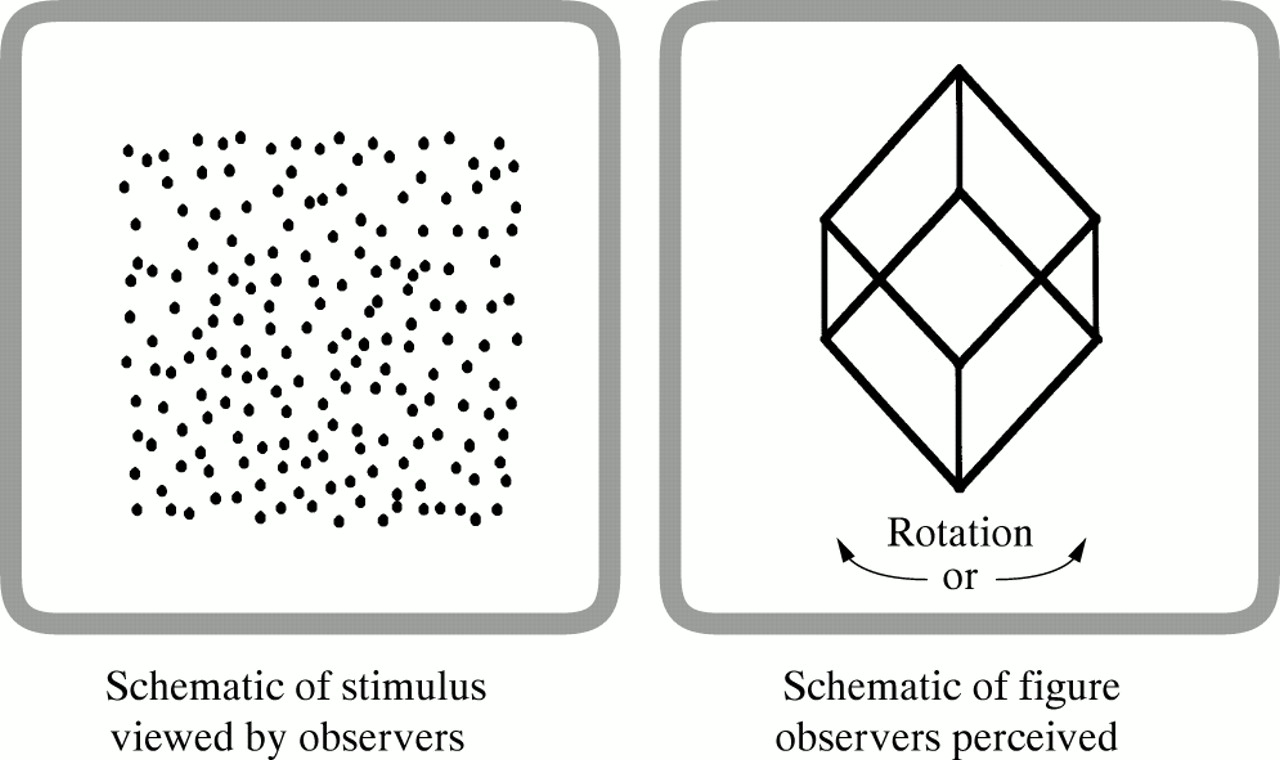
Kinetic depth perception stimulus. The left panel depicts a random dot kinetic depth stimulus with a random dot noise background. Each dot would actually be moving. There is no obvious shape present in the two dimensional structure of the dots. The right panel depicts what is perceived by observers: a three dimensional figure (a cube depicted here) rotating about a vertical axis.
DEPTH FROM MOTION PARALLAX
Previous studies have found that impairments of binocular depth perception (stereopsis) are not sufficient to account for critical features of Bálint's syndrome (for instance, trouble judging the relative depth of objects in space due to an inability to perceive more than one object at a time).10,29 However, other explanations may obtain, including lesions in a system that recovers information of object structure and depth from motion parallax, stereopsis, and other cues.
Knowledge of structure and depth is critical for successful interaction with, acquisition of, and locomotion around objects and obstacles in our environment. This information is so important that the human brain employs a redundant system of multiple visual depth cues including monocular and binocular cues and relative movement. Yet monocular cues to depth (including familiarity, linear perceptive, and interposition) can be deceiving, binocular depth perception (stereopsis) is not available to monocular subjects (or animals with laterally placed eyes), and kinetic depth effect (or structure from motion, see above) may provide ambiguous depth assignments. Motion parallax is one of the most important cues for depth from motion121 and provides unambiguous information on depth from information about direction of subject head movement, and possibly slow eye movements.122
Binocular stereopsis and relative motion may assume greater importance in conditions of high background noise or unfamiliarity. Disparate views from the two eyes generate disparity cues with an orderly geometric relation to relative distance of objects,123 allowing the brain to recover depth information through a neural process resembling triangulation.124 Recovery of depth from relative motion relies on relative movements of retinal images. For motion parallax, relative movement of objects is produced by active movement of the subject, such as moving the head along the interaural axis.125 For motion perspective, relative movement of objects is produced by passive movement of the subject, as when the subject looks out the side window of a moving vehicle.111 In both conditions, objects move in orderly relation to one another across the subject's retina, without changing their location in the world. Objects further than the fixation point seem to move in the same direction the subject's head is moving, whereas closer objects seem to move in the opposite direction. Relative velocities of these movements across the retina have an orderly relation to relative distances of objects in the scene.111,126,127 Such orderly geometric relation make both stereopsis and motion parallax unambiguous cues to relative depth. Unlike motion parallax, structure from motion generates ambiguous depth128,129 and can occur with a stationary subject and relative movement of objects in a scene.128,130,131 Understanding the rules governing combination of different depth cues (stereopsis, motional parallax, structure from motion) into a single percept is an active research topic.132–,134 Feedback signals from slow eye movements may also be important.122 The cerebral lesions associated with Bálint's syndrome are likely to disturb this sytem at multiple levels. Cerebellar lesions may also be implicated. The cerebellum plays a role in integration of head movement, eye movement, vestibular and visual information, in the corticopontocerebellar system for slow eye movement control; and, possibly, in depth from motion parallax. Abnormal perception of depth from motion parallax could accompany smooth pursuit eye moment deficits due to posterior (for example, parietal) hemispheric lesions135,136 or cerebellar lesions, both of which were identified in Bálint's syndrome patient RF49 and in akinetopsia patient LM.119
MENTAL ROTATION
Mental rotation represents a type of visual motion task thought to rely on mental manipulation or movement of an internal representation of a visual stimulus rather than perception of visual movement and is likely to have been impaired in Bálint's patient. In a standard mental rotation task,137 a subject presumably uses mental imagery to rotate a visual stimulus before it can be compared to a standard. Mental rotation may come into play in circumstances when we must identify objects (or faces) encountered in non-canonical orientations, and could place demands on the neural mechanisms for perception of motion. Although relations between reaction time and orientation of visual test figures have been described,138 the underlying mechanisms are not well understood. Right parietal lesions139 (as in those in patients with hemineglect syndrome and Bálint's syndrome) produced longer reaction times, more errors, and differential accuracy over the angle of rotation, compared with normal subjects and subjects with left hemispheric lesions. A mental rotation task may increase activation in right parietal regions, but the left inferior parietal lobes may also play a part.140 Brodmann's areas 39 and 19, thought to contain the human homologue of the mediotemporal area, was selectively activated in both hemispheres in subjects performing a mental rotation task141
We used a version of a mental rotation task142 to test whether selective damage to these areas would affect mental rotation ability in a patient (SF) who had a limited resection of the anterior right occipital gyrus and the posterior sector of both the inferior and middle temporal gyri.143 This includes regions thought to correspond with primate area V5 (mediotemporal area) on the basis of functional imaging.144–,146 Five days after surgery, SF's mental rotation reaction times increased 380%, whereas reaction times on a control (lexical decision) task increased little. SF's mental rotation reaction time function returned to normal by the 18th day, similar to the transient deficits SF showed in tasks of movement perception. The neural mechanisms for mental rotation and for movement perception seem to depend on a human homologue of area V5 (mediotemporal area), which can be affected in some patients with Bálint's syndrome
AWARENESS OF VISUAL IMPAIRMENTS
Holmes noted that patients with suprageniculate lesions “are frequently unaware of loss of vision . . . and consequently may not refer to it in describing their symptoms.”8 He explained that we all have a body scheme that is “built up of . . .visual, tactile, postural and other sensory impressions. When a part of this scheme or image is injured the corresponding part of the body passes out of consciousness, and the patient is no longer aware of it and does not recognise any disturbance within it.”8
Rarely, a patient seems to be completely blind due to a cerebral lesion yet denies the deficit, a situation described by Anton (1899)147 and later called Anton's syndrome.148,149 Anton's syndrome may share a common mechanism with other common forms of anosognosia, such as denial of left hemiplegia or hemisensory loss in patients with a left hemineglect syndrome after right hemispheric damage22 (which is relevant to Bálint's syndrome). Anosognosia has been attributed to disconnection between the right parietal lobe and thalamus150 and may reflect executive dysfunction in patients with frontal lobe lesions.151
Patients who deny blindness generally have large lesions that extend beyond the occipital lobes in association with multiple cognitive defects outside vision. A potential explanation for some cases of Anton's syndrome is failure to conduct an adequate examination—for example, the patient has residual visual function and really is not blind, or has marked cognitive dysfunction that has not been documented.
Patients who have no experience of the items that they localise or detect in the fields of an area V1 (striate cortex) lesion have been described under the oxymoron “blindsight”.152,153 Such patients may perform above chance levels on simple forced choice detection tasks, and on localisation tasks in which the accuracy of finger pointing or eye movements are taken as the index of perception for targets presented in their scotoma. Weiskrantz expanded the definition of blindsight to include visual capacity in a field defect in the absence of acknowledged awareness.154 This definition lacks the original anatomical specificity and covers forms of non-conscious visual processing that may have nothing to do with lesions of V1. Subsequently, blindsight was applied to visual perception in a neglected hemifield155 (such as in Bálint's patient) and can be taken to include knowledge without awareness in patients with agnosia for faces, although it is unclear what blindsight has to do with knowledge.
Current explanations for blindsight may be relevant to explanations of visual performance in some patients with Bálint's syndrome. These explanations include residual function in visual pathways parallel to the retinogeniculostriate system including retinotectal, retinotectopulvinoextrastriate, and geniculoextrastriate pathways,153,156 residual striate cortex,157,158 or partial damage to cerebral networks serving vision.86,159 Striate cortex might be essential for conscious visual perception160 or, alternatively, projections from extrastriate visual cortex to prefrontal cortex may suffice161; yet hemidecorticated patients may retain some visual awareness, raising questions about both these mechanisms.162 The finding of residual vision in blindsight (and in purported cases of Anton's syndrome) could be due to test artifacts, such as inadequate fixation, light scatter, non-visual cues, and non-random presentation of targets.157 The unawareness of the target's presence distinguishes blindsight from residual vision. Those patients retain some awareness of the target within a supposedly dense visual field.163–,166 Vision without awareness also occurs in normal subjects.167,168
VISUALLY GUIDED REACHING AND OPTIC ATAXIA
Bálint's observations on optic ataxia1 sparked interest in the neural basis of visually guided reaching and grasping (fig 6⇓). Reaching and grasping external objects is a fundamental activity that demands the coordination of several different nervous system functions.95,169 To accomplish this task, the brain transforms a target's visual coordinates to body centred space, plans a hand path and trajectory (the sequence of hand position and velocity to target), and computes multiple joint torques, especially about the shoulder and elbow. It also specifies the necessary limb segment orientations from among many possibilities, and activates appropriate muscle groups and inhibits others to meet those specifications. The sensory feedback, frames of reference and neural mechanisms used to solve these complex motor control problems are active research topics.170

Demonstration of optic ataxia in a patient with Bálint's syndrome. The patient is unable to point to the tip of the examiner's finger and grossly overreaches (courtesy of Professor H Stanley Thompson, Professor Emeritus, Ophthalmology, University of Iowa).
Reaching can be divided into different phases. In a transport phase of reaching, the hand is moved toward an object whose position is determined by vision or memory. In an acquisition phase, grasp formation depends on somatosensory and visual information on the limb and target,171 familiarity with the target and, perhaps, predetermined motor programmes. These phases mature at different rates, may be controlled independently before becoming coordinated,172 and can be dissociated by focal brain lesions.94,96,99,173 Posterior parietal damage may affect neurons coding eye position in the head and stimulus location on the retina, which together with neurons in motor and premotor cortex permit hand movements to visual targets in a body centred coordinate system.170 The schema proposed by Soechting and Flanders174–,176 takes origin at the shoulder, is closely related to the representation of object position and motion, and should depend on the visual cortex and its connections to other sensory and motor maps.177–,179
Inability to reach and grasp targets in cases of Bálint's syndrome is often multifactorial, including V1-type visual field defect, defective visual attention, inability to locate targets with the eyes, and abnormal sensorimotor transformations. A range of defects is possible, depending on the extent of a patient's lesions. It seems that cerebral mechanisms for controlling pointing, reaching, and grasping depend on the inferior parietal lobule, temporoparietaloccipital junction, occipital-temporal cortices (particularly areas 37 and 21), and white matter connections. Current evidence suggests that: (1) separate systems exist within these regions to guide distance and direction of reaching36,180; (2) if lesions of the occipital lobe do not extend into areas that project into dorsal stream pathways to the posterior parietal cortex, they do not affect sensorimotor transformations for reaching36,180; (3) superior and inferior parietal lobule lesions and lesions involving the dorsal visual pathways cause abnormally large sensorimotor transformation errors36,181; and (4) mechanisms that control reaching are disturbed by V1 type visual dysfunction only if head and eye movements are constrained.180,182
Relevant to discussions of attention of Bálint's syndrome, it is possible that subjects with lesions in the dorsal visual pathway will have greater difficulty specifying target location when distracters are present. Some types of distracters affect normal subjects' path and accuracy of reaching toward visual targets.183 Distracters may strongly influence visual spatial perception and/or movement control in subjects with lesions of dorsolateral visual association cortices because they increase the complexity of the visual environment, increase attention demands, and may interfere with the neural mechanisms underlying perception and movement planning and execution of both reaching and grasping in normal young adults. Distracters presented centrally,184 for example, but not peripherally185 in the visual field can also influence duration and path of a movement in normal controls. By contrast, one study found hand path deviation toward distracters presented in the visual field ipsilateral to, but not contralateral to, a parietal lesion.186 Patterns of optic ataxia in Bálint's syndrome may vary depending on whether a patient is reaching for a single object, or one object among many. Patterns of recovery in optic ataxia have been reported by Goodale et al.99
Note that optic ataxia must sometimes be distinguished from the “alien hand” phenomenon, in which a patient feels estranged from one of his or her hands.187 The alien hand performs movements that seem beyond volitional control, as if driven by an external agent, and patients may try to restrain it using the good limb. The defect is not just limited to reaching under visual guidance. The affected hand may show a grasp reflex and elements of what Denny-Brown referred to as a “magnetic apraxia”188 associated with frontal lobe damage.189 Optic ataxia may occur as part of Bálint's syndrome in Alzheimer's disease, whereas the alien hand syndrome is more characteristic of cortical basal ganglionic degeneration. Characteristics of cortical basal ganglionic degeneration in 15 patients included “alien limb” phenomena, apraxia, cortical sensory loss, focal reflex myoclonus, rigidity and akinesia, postural action tremor, limb dystonia, hyperreflexia, and postural instability.190 Symptoms and signs are often asymmetric, with greater radiographic evidence of abnormalities in the hemisphere contralateral to the more affected limb. Neuropathological analyses may show swollen, poorly staining (achromatic) neurons and degeneration of the cerebral cortex and substantia nigra, and loss of striatal dopamine. By contrast, patients with Bálint's syndrome and optic ataxia show histopathological changes compatible with Alzheimer's disease.23,24
Mechanisms of optic ataxia in patients with Bálint's syndrome could overlap those of magnetic misreaching, as in “Ms D” who had progressive parietal lobe degeneration and reached “slavishly” toward objects in central fixation when directed to reach toward targets eccentric to fixation.191 Impaired reaching behaviours in Bálint's syndrome may reflect the interaction among different hypothetical parietal lobe and frontal lobe mechanisms for vision and action.192 Lesion of the right frontal lobe may impair monitoring of information related to self generated arm movements.193 The parietal cortex may sustain attention to stimuli in peripheral vision whereas the frontal lobe mediates selective attention through habituation to peripheral stimuli,194 which may explain the fading of visual targets (the Troxler effect) experienced by patients with parietal lobe lesions as opposed to frontal lobe lesions.193 Frontal lobe lesions may impair the suppression of reflexive glances to peripheral stimuli (tested with an “anti saccade” task) and initiation of voluntary glances.195 The frontal lobes may aid in eliminating unwanted reflexive glances and in triggering new movements when the target is not yet known or visible—that is, exerts control over a visual grasp reflex.195
CLINICAL APPROACH
Holmes outlined a scheme for clinical investigations of visual functions of the nervous system that continues to be useful.8 He indicated that visual disorientation can be readily detected, but that it is critical to exclude visual field defects that might interfere with the interpretation. He recommended testing acuity of central vision after errors of refraction had been corrected and opacities and other abnormalities of the media have been allowed for. These days, contrast sensitivity can also be tested (for example, with the Pelli-Robson wall chart). Holmes recommended screening the extent of visual fields for white and colours, firstly by confrontation methods and, if needed, by using a perimeter (to document the presence or absence of visual field suspected by confrontation). He also recommended ophthalmoscopic evaluation, noting the edges and central pit of each optic disc, the colour of each disc, the condition of the retinae, particularly of the macular regions, and the state of the retinal vessels.
Holmes noted that disturbances of visual disorientation can be detected by asking the patient to touch or point in the direction of any objects within the range of vision and to estimate the distance from him by sight alone. He wrote: “Relative localisation is tested by describing the relative positions in space of two objects; when it is defective, he is unable to count correctly dots on a sheet of paper or coins scattered on a table, and he usually fails to divide a line accurately or find the centre of a circle. If able to walk he should be required to find his way about a room in which various obstacles, as chairs or tables, are placed, or through a house, and to describe a route with which he is familiar. On attempting to read he often fails to follow the lines, and especially to bring his eyes to the left of the succeeding line. Unilateral loss of orientation produces less obvious symptoms and is usually not recognised by the subject, he is unable to point to or otherwise indicate, the position of an object in the affected homonymous halves of the fields.”8 Some modern laboratories are equipped to use computerised psychophysical tests of attention and perception; quantitative assessment of hand and eye movements generally requires special equipment and expertise. Current assessment would add relevant standardised neuropsychological tests of visuoperceptual function, as in the case below.
A 70 year old right handed woman complained of increasing visual difficulties for 2–3 years. The patient had fallen on several occasions, and sometimes failed to recognise familiar surroundings and became anxious and tearful. Several ophthalmological assessments showed no visual field deficits and insufficient evidence of eye disease to account for vision complaints. The patient had increasing difficulty with daily activities. For example, after talking on the telephone she had trouble hanging up the receiver on its cradle. She could no longer participate in her lifelong hobbies of sewing and oil painting of pictures because her left arm was becoming difficult to control and sometimes seemed to go off on its own. She had increasing difficulty following conversations and performing multistep commands. The patient had a history of hypertension and hypothyroidism, but no stroke, seizure, or head trauma. She had had a cholecystectomy, hysterectomy, and unilateral oophorectomy. The patient's mother had experienced depression, but there was no dementia, movement disorder, or any other known neurological illness in the family. She had never obtained a driver's licence.
Examination showed an alert, socially appropriate older woman who appeared anxious but not acutely ill. She was aware of and upset by her difficulties. She was oriented to month, city, and person, but not to the year or day of the week. She had striking difficulty with visuomotor tasks, such as copying a simple line drawing of a geometrical figure. Speech was fluent, but she had difficulty following two or more step commands. She scored 16/30 on the Mini Mental State Examination, compatible with dementia of moderate severity. Cranial nerves were intact except for slight limitation of upgaze. She had good muscle strength, but complained that her left hand wouldn't do what she wanted it to do. On one occasion her hand had seemed to “levitate” by her side. She had difficulty with skilled limb movements, especially with the left. She had mild left hand dystonia. Visually guided reaching was severely impaired, especially on the left, but there was also some difficulty reaching to targets on her own body. Gait was slow and cautious, with reduced arm swing. She had difficulty locating and pointing towards a mirror on a door in the examining room. While supine, upon command to raise her left leg, there was a delay of a few seconds before she raised her right leg. Pinprick and touch were intact. Pallesthesia and proprioception were normal in the hand and feet. Tendon stretch reflexes were graded as 2/2 and symmetric. Babinski's sign was absent. There was no myoclonus or tremor. Haematological studies showed normal blood counts, thyroid functions, B12 and folate, liver function tests, and a rapid plasma reagin test was non-reactive.
Neuropsychological testing disclosed severe deficits on all tests involving ocular motor, visuospatial, and visuomotor abilities, including tests of constructional praxis, gestural praxis, visual scanning, manual dexterity, and visually guided reaching (fig 7⇓). The patient was unable to give a coherent report of the Cookie Theft picture. She also had significant impairments on tests of orientation, verbal memory, working memory, and speech and language (controlled word finding, aural comprehension, and confrontation naming). Electroencephalography showed slowing of the background rhythms in both hemispheres, without epileptiform discharges or triphasic waves. Brain MRI was read as within normal limits for age. However, PET showed marked hypometabolism of both parietal lobes (right greater than left), in the superior portion of both temporal lobes (right greater than left), and the lateral and midline portion of both occipital lobes. The basal ganglia and frontal lobes were intact. Transverse, coronal, and sagittal PET images are shown in figure 8⇓.

Top panel: abnormal pointing movements are shown from a patient with features of Bálint's syndrome, in the context of the visual variant of Alzheimer's disease. The overhead view shows the paths of multiple pointing movements between a target directly in front of her body, in the midline, to more distal targets located to the left, middle, and right. Multiple pointing movements are shown to each target with the left hand (left top panel) and right hand (right top panel). The patient made highly erratic movements (especially with the left hand). Bottom panel: in comparison, a subject of similar age without neurological disease made smooth pointing movements. The recordings were made using an optoelectronic technique to track infrared emitting diodes on the patient's index finger.

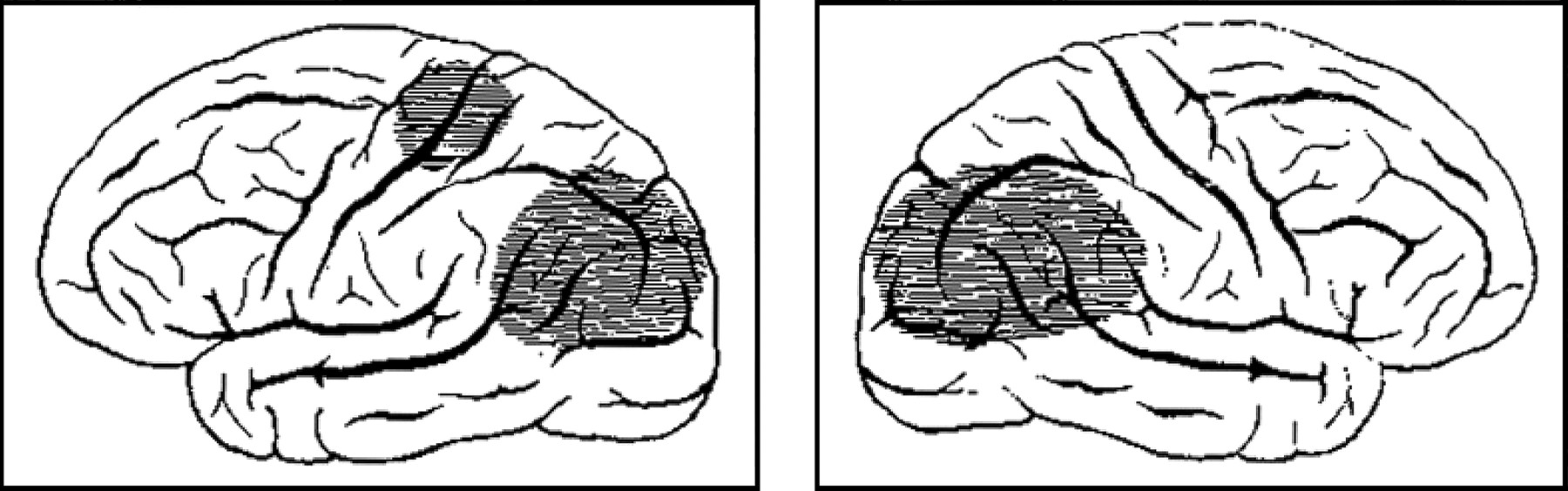{kind=link}
{kind=link}
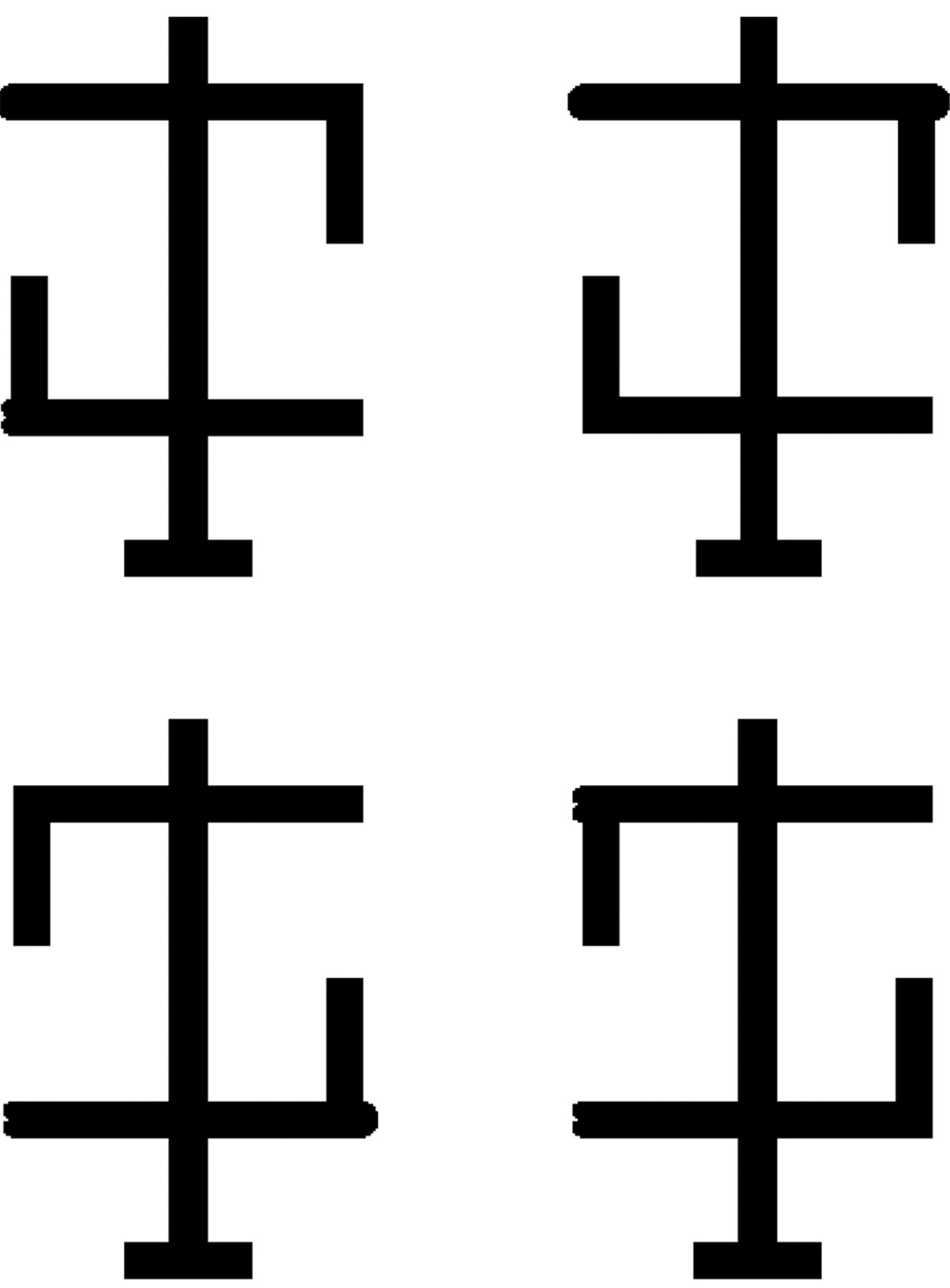{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
FDG PET in the same patient with Bálint's syndrome whose handpaths are depicted in Figure 7⇑. (A) Transverse view; (B) coronal view; (C) sagittal view. The blue colour in all pictures indicates abnormally low cerebral activity, especially in the parietal regions.
The main features in this patient are insidious onset of progressive difficulties with vision, reaching, and cognition and biparietal hypometabolism. This picture is broadly compatible with Bálint's syndrome due to Alzheimer's disease. The left hand reaching difficulty resembled optic ataxia, but atypical features included mild left hand dystonia with reported “wandering” of the left hand. These are features of the “alien hand syndrome” seen in patients with parietal lobe lesions due to stroke and in cortical basal ganglionic degeneration. The absence of myoclonus and of triphasic waves in the EEG fails to provide support for spongiform encephalopathy (Creutzfeld-Jakob disease). Mild limitation of upgaze, mild left hand dystonia, and reduced left arm swing suggest a basal ganglionic component, as in Lewy body disease, but the basal ganglia were normal on PET imaging. There was little support for a systemic illness or an opportunistic infection such as progressive multifocal leukoencephalopathy, which often affects posterior white matter and should have been visible on the MR images. Three years after presentation, the patient's abilities had declined along several cognitive domains, her visual impairments were part of a broader picture of severe dementia, and she was unable to take responsibility for most of her personal needs.
CONCLUSIONS
Visual cues convey a wealth of information on physical objects and their relations. We construct our reality from serial glimpses, and our impression of a seamless and richly detailed visual world is an illusion. Damage to visual cortex and white matter in Bálint's syndrome can destroy this illusion, resulting in a piecemeal experience of the visual world (simultanagnosia/visual disorientation) and impaired visual control of eye movements (ocular apraxia) and hand movements (optic ataxia). Evidence from patients who display elements of Bálint's syndrome is relevant to the understanding of the neural substrates of visual perception, motor control, attention, working memory, and even consciousness. Although Bálint's syndrome has been construed as a biparietal syndrome causing an inability to see more than one object at a time; other lesions and mechanisms are possible. Key syndrome components are dissociable and comprise a range of disturbances that overlap the hemineglect syndrome. As Bálint's syndrome is infrequent and difficult to assess with standard clinical tools, the literature is dominated by case reports; however, it is risky to generalise from single case reports in Bálint's syndrome. Future studies should address possible underlying psychoanatomical mechanisms at “bottom up” and “top down” levels, with specific consideration to visual working memory and attention (including object attention), and to systems for identification of object structure and depth from binocular stereopsis, kinetic depth, motion parallax, eye movement signals, and other cues.
Acknowledgments
This work was supported by NIH-NINDS PO1 NS 19632 (anatomical basis of vision) and NIH-NIA RO1 AG/NS 15071 (driving in old age, Alzheimer's disease, and stroke)